


{kind=link}
{kind=link}
{kind=link}
{kind=link}
北京市3个公园白纹伊蚊种群 Rdl抗性基因突变分析
[周小洁1 , 张洪江2 , 赵显丰3 , 邱星辉4 , 张勇1, *  ]
]
]
|
|


作者简介:周小洁(1981—),男,博士,副研究员,研究方向:媒介生物控制研究。
目的 对北京市3个大型森林公园白纹伊蚊( Aedes albopictus)野外种群的 Rdl基因抗性突变进行检测,评估其对γ-氨基丁酸(γ-aminobutyric acid,GABA)受体类杀虫剂的抗性风险。方法 利用序列比对分析获得白纹伊蚊编码GABA受体的Rdl基因序列,通过PCR扩增 Rdl基因片段并进行测序,分析与抗性有关位点(第A296和V327位点)的突变情况。结果 白纹伊蚊 Rdl基因由10个外显子(Exon)和9个内含子(Intron)组成,编码区(CDS)全长1 608 bp,编码535个氨基酸;白纹伊蚊 Rdl基因编码氨基酸序列与埃及伊蚊( Aedes aegypti)和淡色库蚊( Culex pipiens pallens)相应序列的相似性分别为99.63%,与黑腹果蝇( Drosophila melanogaster)相似性为91.55%;在3个公园采集野外白纹伊蚊种群中(90只标本)均未检测到上述 Rdl靶标抗性突变,均为敏感型(GCA和GTA)。结论 北京市3个大型的城市森林公园环境中白纹伊蚊种群对环戊二烯类有机氯杀虫剂和苯基吡唑类杀虫剂氟虫腈的靶标(即GABA受体) Rdl基因与其他昆虫表现出不同程度的同源性和高度保守的基因结构特征,3个受试白纹伊蚊野外种群未检测到 Rdl基因抗性相关突变,提示以靶标敏感为导向反向研发杀虫剂。
Objective To analyze Rdl resistance mutations in three park-collected populations of Aedes albopictus in Beijing, and assess their risk of resistance to cyclopentadienyl insecticides.Methods Rdl gene sequence encoding the γ-aminobutyric acid (GABA) receptor in A. albopictus was obtained by sequence retrieve and alignment analysis, then amplified by PCR and sequenced to analyze mutations at resistance-related loci (A296 and V327).Results Rdl gene of A. albopictus consists of 10 exons and 9 introns, with a coding region (CDS) of 1 608 bp in length, it codes for 535 amino acids; The amino acid sequence of the Rdl gene of A. albopictus showed 99.63% similarity to the corresponding sequences of A. aegypti, Culex pipiens pallens , and 91.55% similarity to Drosophila melanogaster. No mutation was detected in the above loci in three field populations of A. albopictus (90 specimens), all were sensitive genotype (GCA and GTA).Conclusions The study was conducted in three large populations of A. albopictus in urban forest park environments in Beijing, and investigated the gene characteristics and resistance mutations of the targets (GABA receptors) of cyclopentadiene organochlorine insecticides and the phenyl pyrazole insecticide fipronil, and found that the Rdl gene of A. albopictus showed different degrees of homology and highly conserved gene structure characteristics with other insects, and no Rdl resistance-related mutation was detected in three field populations of A. albopictus, suggesting that target-sensitive-oriented reverse development of insecticides.
白纹伊蚊(Aedes albopictus)是基孔肯雅病毒、登革热病毒、黄热病病毒和寨卡病毒[1]等虫媒病毒的主要传播媒介。白纹伊蚊适应性强, 呈现全球扩散趋势, 在我国大部分地区均有分布, 主要依靠孳生地治理和化学杀虫剂进行防制。白纹伊蚊作为北京市传病风险较高、叮咬危害严重的第二优势蚊种[2], 对其生态学、抗药性和病原学监测是有效防制媒传疾病的基础。有报道[3], 对蚊虫进行化学防治可造成的持续选择压力, 将导致蚊虫对杀虫剂产生抗性。蚊虫对杀虫剂的抗性机理主要包括解毒作用增强和靶标不敏感[4], 前者主要与酯酶、细胞色素P450氧化酶、谷胱甘肽S-转移酶等解毒酶活性增加有关, 后者则与钠离子通道、乙酰胆碱酯酶、γ -氨基丁酸(GABA)受体等杀虫剂靶标突变进而降低与杀虫剂的亲和力有关。本研究主要针对环戊二烯类杀虫剂靶标GABA受体在三个白纹伊蚊野外种群中的突变情况开展研究。
1.1.1 试虫采集 于2018年8— 9月在北京市朝阳区奥林匹克森林公园(OFP)、石景山松林公园(SLP)和通州大运河森林公园(TCP), 同时用二氧化碳诱蚊灯和人诱停落法在下午4∶ 00~7∶ 00进行诱集白纹伊蚊, 经形态学鉴定后将其浸于75%乙醇溶液, 于4 ℃保存直至后续分析。从采集到3个白纹伊蚊野外种群中各取30只雌蚊用于后续试验分析。
1.1.2 试剂与仪器 蛋白酶K、T-载体购自Promega, 基因扩增检测所需2× EZ1 PLUS Taq PCR MasterMix(Red)与DNA marker来自北京达科为生物技术有限公司。PCR扩增仪、离心机与电泳设备来自日本ASTEC, 核酸微量测定仪(ND-1000)来自美国NanoDrop。引物合成与基因测序由北京擎科新业生物技术有限公司提供。
1.2.1 序列获取与比对 从NCBI的白纹伊蚊基因组数据库获取Rdl基因序列信息, 从GenBank数据库中获取埃及伊蚊(Aedes aegypti)的Rdl基因序列, 将二者的DNA序列及编码序列进行比较, 进而确定白纹伊蚊Rdl基因结构(外显子-内含子构成)。用MUSCLE(3.8)软件中CLUSTAL程序对来自果蝇(Drosophila melanogaster)、埃及伊蚊(A.aegypti)和白纹伊蚊的Rdl基因氨基酸序列进行多序列比对。
1.2.2 基因组提取 参考Rinkevichet al.的方法[5], 提取单只白纹伊蚊(去除腹部)的基因组DNA, 用NanoDrop测定基因组样品的DNA浓度和提取质量。
1.2.3 基因扩增与测序 以单只白纹伊蚊基因组DNA为模板, 采用引物组合AaRDL-F(cagtttgtacgttcgatgggt)和AaRDL-R(ggcaaataccatgacgaagca)扩增编码GABA受体β 亚基的Rdl基因含上述位点的基因片段。设置2个阴性对照, 一个是实验室敏感种群白纹伊蚊基因组作为模板进行扩增, 另一个为不加基因组DNA的空白反应体系。PCR反应体系为30 μ L, 包含15 μ L的2× Taq PCR MasterMix, 正、反向引物(终浓度10 μ Mol), DNA模板100~200 ng, 去离子水补齐至 30 μ L。PCR反应条件:94 ℃预变性5 min; 94 ℃变性10 s, 60 ℃复性30 s, 72 ℃延伸35 s, 共38个循环; 最后72 ℃延伸10 min。上述PCR产物经2.0%的琼脂糖凝胶电泳检测, 确认扩增成功后用正向引物AaRDL-F进行测序。
1.2.4 基因突变分析 对于目标扩增片段, 将每个种群的30个扩增片段中第296和327位点的编码氨基酸与数据库中的序列进行比对, 分别计算不同种群上述位点的突变频率。
从埃及伊蚊基因组信息中检索其Rdl基因DNA序列, 其Gene ID是U28803.1, 根据该基因编码的氨基酸序列推导出白纹伊蚊Rdl基因(Gene ID:109622845)的结构及序列, 发现其由10个外显子(Exon)和9个内含子(Intron)组成, 编码区(CDS)全长1 608 bp(见表1), 二者基因结构保持一致。将果蝇(D. melanogaster)、埃及伊蚊(A.aegypti)和白纹伊蚊(A.albopictus)的Rdl基因编码的氨基酸序列进行比对, 三者之间序列相似性较高(见图1), 尤其是埃及伊蚊和白纹伊蚊序列相似性高达99.63%, 与黑腹果蝇(D. melanogaster)Rdl编码序列相似性为91.55%。
| 表1 白纹伊蚊Rdl基因(Gene ID:109622845)结构 Table 1 Genomic structure of Rdl gene in A. albopictus |
 | 图1 不同物种Rdl基因编码氨基酸序列比对 ∗.表示该位点氨基酸在3个比对序列间完全保守, ∶ .表示该位点的氨基酸残基较为保守, ▪.表示该位点的氨基酸残基保守性较差。红色标记的296和327位点代表本研究分析的2个抗性突变位点。Fig. 1 Comparison of amino acid sequences encoded by Rdl genes of different species ∗. Indicate identical sites among the 3 sequences; ∶ .Represent sites with conserved substitutions, and ▪. Stand for sites with weakly conserved sites in the 3-sequence alignment. The 296 and 327 sites marked in red indicate the 2 resistance mutations of interest in this study. |
对目标基因进行扩增, 将扩增成功的产物进行基因测序, 扩增条带符合预期产物长度246 bp, 见图2。阴性对照敏感种群白纹伊蚊可正常扩增, 空白对照不加基因组模板无扩增产物。
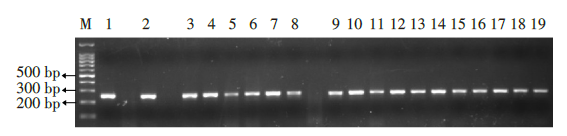 | 图2 扩增Rdl基因片段电泳结果。 M.100 bp marker, 第1~9泳道为OFP种群扩增产物, 第10~19泳道为TCP种群扩增产物, 扩增产物长度为246 bp。Fig. 2 The results of amplified Rdl gene fragment electrophoresis M. 100 bp marker, lanes 1-9 for OFP stock amplification product, lanes 10-19 for TCP stock amplification product, amplification product length is 246 bp. |
本研究关注的Rdl基因突变位点, 位于7号外显子内, 从中选择部分目标序列进行扩增, 扩增片段包含了296和327位点为广泛报道的有意义突变位点(见图3中绿色部分)。对三个种群白纹伊蚊Rdl基因的扩增片段测序发现, 296位密码子均为敏感型GCA(A296), 327位密码子也均为敏感型GTA(V327), 未检测到抗性突变情况(见图4)。
 | 图3 本研究中扩增的白纹伊蚊Rdl基因片段位于第7号外显子 黑色圆点(●)表示与抗性相关的2个突变所在氨基酸位点(296和327)。Fig. 3 The A. albopictus Rdl gene fragment amplified in this study located in exon 7 The dots (●) indicate the amino acid sites (296 and 327) related with resistance mutation. |
 | 图4 三个野外种群(OFP、SLP、TCP)白纹伊蚊Rdl基因代表性扩增片段中296和327位密码子突变情况Fig. 4 Mutation of codons 296 and 327 in the representative amplified fragment of the Rdl gene of three field populations (OFP, SLP, TCP) of A. albopictus |
白纹伊蚊是基孔肯雅热、登革热、黄热病和寨卡病毒等疾病的重要传播媒介。在缺乏特异性抗病毒药物和商业化疫苗的情况下, 蚊虫防制仍然是防控该类传染病的最有效手段。而化学防治又是蚊虫防制不可或缺的手段, 但随之也带来了不可避免的抗药性问题。
GABA受体是多氯环烷烃类杀虫剂(如硫丹、艾氏剂、狄氏剂等环戊二烯类, 以及林丹等六氯环己烷异构体)和苯基吡唑类杀虫剂(如氟虫腈)的作用靶标。本研究对北京地区3个代表性大型城市森林公园野外白纹伊蚊种群中GABA受体编码基因Rdl分子特征及抗性突变进行研究。通过数据库检索、序列分析比对获得白纹伊蚊Rdl基因序列, 分析其基因结构, 其表现出与埃及伊蚊和黑腹果蝇Rdl完全相同的基因结构, 与埃及伊蚊Rdl编码区的相似性高达99.63%, 表明具有高度的同源性。在此基础上, 对文献中确认的Rdl基因抗性突变信息进行分析, Michael等(1993)从埃及伊蚊中发现Rdl基因突变(A302S), 该突变同样存在果蝇中, 且经功能试验证实其与抗性密切相关[6]。同时, 该突变还在家蝇[7]、蟑螂[8, 9]、中华按蚊[10]等卫生害虫以及小菜蛾[11]等农业害虫中存在。
截至2022年, 关于白纹伊蚊Rdl抗性突变研究报道仅有2篇, 分别是来自留尼汪岛[12]和马来西亚[13]的白纹伊蚊种群。在印度洋中的留尼汪岛, 尽管环戊二烯类杀虫剂禁用已超15年之久, 全岛多种蚊虫种群内仍有很高的Rdl抗性突变频率, 并在两个白纹伊蚊的样本中首次检测到该抗性等位基因, 突变频率分别为10%和30%。本研究对北京市3个不同方位的大型城市森林公园采集的白纹伊蚊种群进行检测, 结果未检测到Rdl抗性突变, 原因可能与本地的用药历史、伊蚊时空分布与自然环境变化有关。2001年5月23日, 包括中国在内的127个国家和地区在瑞典首都斯德哥尔摩签署了一项国际公约, 决定在世界各地禁止或限制使用12种持久性有机污染物, 其中就包括上述环戊二烯类杀虫剂[14]。本研究采样3个公园在建成前所处的环境为农村用地, 可能有环戊二烯类杀虫剂应用历史, 但北京市农业生产主要集中在远郊区, 杀虫剂使用强度较低, 同时该类杀虫剂较早就被禁用, 对蚊虫的选择压力未能持续形成; 此外, 另一原因是北京市白纹伊蚊的种群规模仅在近五年有较大提高, 这与气候变暖、经济发展导致人流贸易增多、城镇化增加有关, 而在此之前白纹伊蚊占比较低[15], 同时白纹伊蚊以小型积水为主要孳生环境有关, 接触该类杀虫剂的机率较低。综上所述, 基于采样环境用药历史、伊蚊种群占比及其生态习性等因素, 导致在3个公园环境采集的白纹伊蚊种群中未检测到Rdl抗性基因突变。对于苯基吡唑类杀虫剂氟虫腈, 因其只能用于卫生杀虫剂, 主要用于室内蟑螂、白蚁等爬虫的防制, 与白纹伊蚊生态位无明显重合, 无明显选择压力。本研究中未检测到白纹伊蚊种群发生Rdl基因抗性突变, 但作为蚊虫防制的基础工作, 对不同杀虫剂的抗性水平以及机理的监测研究是必不可少的, 同时也提示可以针对蚊虫该靶标反向进行新杀虫剂种类研发。
利益冲突声明 所有作者声明不存在利益冲突
编辑:符式刚
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|